Disponível
em:
http://www.fcagr.unr.edu.ar/malezas/Apuntes/Funciones_Dano.doc
FUNCIONES DE DAÑO DE MALEZAS
![]()
Dr. Javier I. Vitta. Cátedra de Malezas. Facultad de
Ciencias Agrarias. Universidad Nacional de Rosario. CC14. 2123 Zavalla.
Argentina.
![]()
Uno de los objetivos en el manejo de
malezas es poder predecir las pérdidas por competencia. Diferentes tipos de
modelos han sido ideados con esa finalidad. Los mismos abarcan distintos
niveles de elaboración: desde modelos sencillos de regresión hasta complejos
modelos ecofisiológicos, los cuales identifican los recursos por los cuales se
compite. En este trabajo abordaremos el primer tipo de modelos: describiremos
los tipos más comunes, analizando sus
ventajas y limitantes.
Las funciones de daño se caracterizan
por ser modelos descriptivos, es decir, que no consideran los factores
involucrados en el proceso de competencia. Son principalmente generados
mediante diseños experimentales aditivos, combinando una única densidad del
cultivo con diferentes densidades de la maleza. Los modelos así obtenidos
consisten, por lo general, en regresiones simples que describen la pérdida de
rendimiento den función de distintas variables que caracterizan la abundancia
de la maleza. Entre ellas, la biomasa es desde un punto de vista biológico, la
más adecuada para cuantificar la demanda de recursos (luz, agua y nutrientes),
y por ende, la intensidad de la competencia entre cultivos y malezas. Su empleo
con fines predictivos se ve seriamente restringido debido a que su determinación
supone un muestreo destructivo, además de ser engorrosa desde un punto de vista
operativo. No obstante, es utilizada con fines experimentales para analizar la
complementariedad de recursos entre cultivos y malezas y/o la agresividad
relativa de ambas especies.
Históricamente, los modelos estáticos más utilizados consideran la
densidad de la maleza como variable independiente. Esta variable es más fácil
de determinar que la biomasa y permite establecer relaciones estadísticamente
aceptables con el rendimiento del cultivo, incluso desde etapas tempranas del
ciclo de crecimiento. Numerosos
trabajos han sido llevados a cabo con el objeto de describir la forma de la
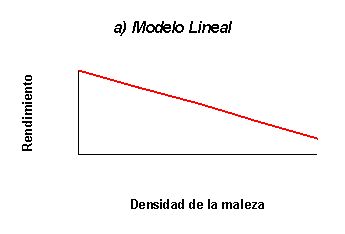
función que relaciona ambas variables. Así, por ejemplo, Dew (1972) estableció
una relación lineal entre la pérdida de rendimiento de distintos cultivos y la
raíz cuadrada de la densidad de plantas; Torner et al. (1991) optaron por una
relación exponencial para describir el rendimiento de cebada en función de la
densidad de Avena sterilis; Leguizamón
y Vitta (1989) ajustaron un modelo lineal que relacionó el rendimiento de soja
en función de la densidad de macollos de Sorghum
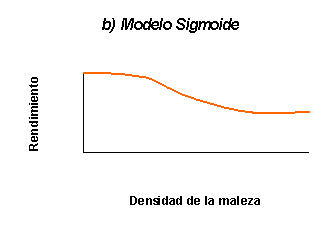
halepense. Zimdahl (1980), por su parte propuso una relación general de
tipo sigmoide entre la disminución del rendimiento de los cultivos y la
densidad de las malezas.
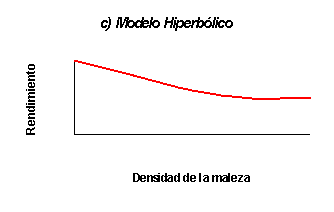
Cousens et al. (1984) sostienen que la elección de los modelos
previamente mencionados usualmente se basa en consideraciones estadísticas, sin
alguna restricción de tipo biológico. Para paliar este déficit, los autores
idearon un modelo hiperbólico cuyos parámetros poseen un significado biológico
concreto:
Pr =
. iD . (1)
1+
(iD/m)
Donde:
Pr= pérdida de rendimiento del
cultivo (g/m2)
D= densidad de la maleza (pl/m2)
i= pérdidas que resultan a
densidades muy bajas de la maleza
m= pérdida máxima de rendimiento
Dicho modelo contrasta con la interpretación biológica que surge de considerar el modelo sigmoide
propuesto por Zimdahl. El modelo hiperbólico supone que en bajas densidades la
competencia interespecífica es máxima. En estas condiciones, un aumento en la
densidad de la maleza tendrá un efecto aditivo sobre el rendimiento del
cultivo. En altas densidades, las plantas de la maleza compiten entre sí de
manera que la relación deja de ser lineal para tornarse curvilínea. Por el
contrario, el modelo sigmoide asume que existe una densidad de la maleza por
debajo de la cual no existe competencia. La interpretación más aceptada en la
actualidad es que el modelo sigmoide deriva de la imposibilidad de detectar
disminuciones estadísticamente significativas cuando las densidades son muy
bajas. El modelo lineal, por otra parte parece ser adecuado para situaciones de
baja densidad de malezas, donde el efecto competitivo de las mismas es aditivo.
En la Figura 1 se describen las principales formas de respuesta
mencionadas.
La principal desventaja de las funciones de daño basadas en la
densidad es su incapacidad para cuantificar los efectos de las malezas en
distintas situaciones de competencia. En
la década pasada, se realizaron distintos intentos considerando otras variables
además de la densidad de la maleza, con la finalidad de aumentar la generalidad
de los modelos mencionado. Así, Carlson y Hill (1985) demostraron que la
inclusión de la densidad de trigo como factor de competencia con Avena
fatua mejora sustancialmente el valor predictivo de la función de daño del
cultivo. Por su parte, Cousens et al. (1987) construyeron un modelo no lineal
incorporando como variable independiente, además de la densidad de la maleza,
el momento relativo de emergencia de la misma.
A pesar de estos intentos, la
varianza no explicada por estos modelos sigue siendo importante. Los parámetros
estimados suelen variar significativamente entre años y entre sitios. Cousens et al. (1988) observaron que el orden de variación en la respuesta del trigo
a la competencia de Bromus sterilis
entre distintas localidades de Inglaterra –atribuido, al menos parte, a
diferencias edáficas entre sitios- desalentaba el empleo de las funciones de
daño como herramienta de manejo de la maleza. Sin embargo, en ciertos casos, la
robustez del modelo puede ser suficiente como para extender sus predicciones a
una escala regional (Gill et al., 1987).
Desde un punto de vista práctico,
la precisión requerida al modelo dependerá tanto del nivel de rendimiento
esperado del cultivo como de la agresividad de la maleza. En la medida que
ambos aumenten , la pérdida deberá ser estimada con mayor exactitud.
Vitta (1994) demostró que el
efecto competitivo de la maleza a bajas densidades (estimado por el parámeto i de la ecuación (1)) es más variable
que la pérdida a muy altas densidades (estimada por el parámetro m de la misma ecuación). Este hecho es
particularmente relevante teniendo en cuenta que la toma de decisiones acerca
del control de malezas es especialmente crítica con bajos niveles de
infestación.
Assemat (1988) señala que la falta
de generalidad de las funciones de daño deriva de la ausencia de un vínculo
funcional entre la densidad de la maleza y el rendimiento del cultivo. Este
último no responde al número medio de individuos competidores, sino a la
demanda de recursos que ellos ejercen. Los modelos basados en la densidad de
las malezas no consideran la heterogeneidad ambiental que gobierna las
relaciones de competencia. Por lo tanto, el valor predictivo de este tiipo de
modelos se restringe a condiciones
ambientales similares a las existentes durante su generación.
Figura 1: Principales formas de las funciones de daño de malezas
Kropff y Spitters (1991)
propusieron considerar el área foliar relativa de la maleza en lugar de su
densidad. Los autores propusieron el siguiente modelo que relaciona las
pérdidas de rendimiento (Pr) y el área foliar relativa de malezas (AFm).
Pr = qAFm
(2)
1+ (q-1) AFm
q es el coeficiente de daño por
competencia. AFm representa la
proporción del área foliar de la mezcla que corresponde a la maleza.
La ecuación (2) asume que el
canopeo es un adecuado integrador de los distintos factores involucrados en la
competencia. En particular, los modelos basados en el área foliar – a
diferencia de los basados en la densidad- tienen en cuenta las variaciones en
el momento relativo de emergencia de las especies: las malezas mas tardías –y
por lo tanto, menos competitivas- representarán una menor fracción del Afm en
comparación a aquellas malezas que emergen simultanemente con el cultivo. Existe
ya evidencia experimental que demuestra la superioridad de esta nueva aproximación.
No obstante ello, el área foliar presenta dos inconvenientes:
i)
lo dificultoso de su
determinación.
ii)
la diferente tasa de crecimiento
del área foliar de las especies. Esto último provoca que el parámetro q de la regresión se modifique según el
momento del ciclo en que se haya determinado el área foliar.
Para contrarrestar la primera
limitante, en los últimos años se han ideado distintas metodologías de
estimación del área foliar de malezas, basadas en evaluaciones de
cobertura. Pike et al. (1990) observaron
que la cobertura de Datura stramonium
y Xanthium strumarium -estimadas
mediante fotografías analizadas con técnicas de imagen- predijo adecuadamente
el rendimiento del cultivo de soja. Ghersa y Martinez Ghersa (1991)
desarrollaron un método para predecir la competencia entre maíz y S. halepense utilizando un medidor
óptico ideado por Beaumer y De Wit (1968). El método fue probado a nivel
extensivo, en donde el rendimiento calculado del cultivo difirió del observado
en no más del 6%. Vitta et al. (1993), comprobaron la bondad predictiva de ese
método en mezclas de soja y S. halepense.
Lutman (1992) observó una alta correlación entre la materia seca de avena, su
cobertura –estimada visualmente o por fotografías- y el rendimiento final de
vicia y arveja. Christensen (1993) trabajó en determinaciones electrónicas de
la cobertura de Brassica napus en el
cultivo de cebada. El método utiliza como variable independiente la
intercepción relativa de luz de la maleza y su empleo parece restringirse a
propósitos experimentales. Finalmente, Vitta y Fernandez Quintanilla (1996)
trabajando con mezclas de Sinapis alba
y trigo, concluyen que estimaciones visuales de la cobertura predijeron en
forma más acertada el rendimiento del cultivo que métodos más objetivos basados
tanto en fotografías como en el uso del medidor óptico antes mencionado.
Las funciones de daño basadas en
la cobertura de malezas pueden ser tanto lineales (v.gr. soja-S.halepense) como hiperbólicas (v.gr.
trigo-S.alba). Si bien estas
funciones en muchos casos explican un alto porcentaje de la variación del
rendimiento de los cultivos, sus parámetros pueden modificarse de acuerdo a
cambios en el ambiente. Por ejemplo, Vitta et al. (1993) encontraron que un
mismo nivel de cobertura de S. halepense
produce una mayor pérdida por competencia en un cultivo de soja de segunda que
en uno de primera.
Existen además otra serie de
restricciones que limitan el uso de las funciones de daño con herramientas de
manejo de malezas. Sumariamente, ellas se basan en las siguientes
consideraciones:
(i)
la distribución espacial de malezas.
El hecho de tener en cuenta el
promedio de la densidad o de la cobertura para caracterizar el nivel de
infestación de una determinada especie, significa asumir que la distribución de
la misma es azarosa. Sin embargo, las malezas normalmente se encuentran en
manchones y se ajustan a una distribución binomial negativa (Marshall, 1988). En
estos casos, el supuesto de distribución al azar provoca una sobreestimación de
las pérdidas reales por competencia cuando estas responden a un modelos
hiperbólico (Brain y Cousens, 1990). Tanto las posibles correlaciones entre las
distribuciones de diferentes especies como la variabilidad registrada en la
distribución entre distintos campos, dificultan la modelización adecuada de la
distribución de malezas (Wiles et al., 1992).
(ii)
el período de tiempo que abarque la evaluación
económica del control.
Por lo general, el umbral de
infestación a partir del cual es económicamente rentable controlar una maleza
es menor cuando el análisis incluye no sólo el año actual sino también los
sucesivos. Por ejemplo, Bauer y
Mortensen (1992) estimaron que el umbral económico a corto plazo de Abutilon theophrasti y Helianthus annuus en soja era 2.6 y 1.5/
10 m2, respectivamente. En cambio, a largo plazo (considerando un
período de 15 años) los umbrales se reducían a 0.4 plantas/10m2 para
ambas malezas. Para realizar este tipo de análisis, es necesario incorporar a
las funciones de daño un modelo de la dinámica de la población de la maleza.
(iii)
la coexistencia de distintas especies de malezas.
Generalmente, las funciones de
daño consideran que las pérdidas atribuidas a la competencia con una única
especie de maleza. Sin embargo, la situación más común es la presencia de
distintas especies compitiendo entre sí y con el cultivo. Distintos aportes
destinados a superar este inconveniente se basan en el cálculo de un índice de
competencia que engloba todas las especies presentes. Dicho índice deriva de
multiplicar la habilidad competitiva de cada especie por su densidad (Coble y
Mortensen, 1992). En el caso de los modelos basados en área foliar (ecuación
(2)), la alternativa es realizar la sumatoria del area foliar relativa de cada
especie de maleza por su respectivo coeficiente de daño (q).La limitante de
estas aproximaciones es la incapacidad de cuantificar adecuadamente la
competencia intra e interespecífica de las malezas.
Bibliografía
·
Assemat, L. (1988).
Proceedings of the 8th. Symposium of Weed Biology, Ecology
and Systematics, Dijon, 649-656.
·
Baeumer, D. y De Wit, C.T. (1968).
Netherlands
Journal of Agricultural Science, 16,
103-122.
·
Bauer, T.A. y Mortensen, D.A.
(1992). Weed Technology, 6, 228-235.
·
Brain, P. y Cousens, R. (1990). Journal of Applied Ecology, 27, 735-742.
·
Carlson, H.L. y Hill, C.T. (1985). Weed Research, 33, 176-181.
·
Coble, H.D. y Mortensen, D.A.
(1992). Weed Technology, 6, 191-195.
·
Cousens, R., Peters, N.C.B. y
Marshall, C.J. (1984). Proceedings of the
7th. Symposium of Weed Biology, Ecology and Systematics, Dijon,
367-374.
·
Cousens, R., Brian, P., O’Donovan,
J.T. y O’Sullivan, A. (1987). Weed Science, 35, 720-725.
·
Cousens, R., Firbank, L.G.,
Mortimer, A.M. y Smith, R.G.R. (1988). Journal
of Applied Ecology, 25, 1033-1044.
·
Christensen, S. (1993). Proceedings of the EWRS Symposium,
Braunshweig, 63-70.
·
Dew, D.A. (1972). Canadian Journal of Plant Science, 52,
921-927.
·
Ghersa, C.M. y Martinez Ghersa, M.A.
(1991). Weed Technology, 5, 279-285.
·
Gill, G.S., Poole, M.L. y Holmes,
J.E. (1987). Australian Journal of
Experimental Agriculture, 27, 291-294.
·
Kropff, M.J. y Spitters, C.J.T.
(1991). Weed Research, 31, 97-105.
·
Leguizamón, E.S. y Vitta, J.I.
(1989). Actas de la IV Conferencia
Mundial de Investigación en Soja, Buenos Aires, 1632-1638.
·
Lutman, P.J.W. (1992). Proceedings of the 9th. Symposium
of Weed Biology, Ecology and Systematics, Dijon, 337-344.
·
Marshall, E.J.P. (1988). Weed Research, 28, 191-198.
·
Pike, D.R., Stoller, E.W. y Wax,
L.M. (1990). Weed Science, 38,
522-527.
·
Torner, C., Gonzalez Andújar, J.L.,
Fernandez Quintanilla, C. (1991). Weed
Research, 31, 301-307.
·
Vitta, J.I., Satorre, E.H. y
Leguizamón, E.S. (1993). Weed Research, 34, 89-97.
·
Vitta, J.I. (1994). Tesis
Doctoral-Universidad Politécnica de Madrid. 150 pp.
·
Vitta, J.I. y Fernandez
Quintanilla, C. (1996). Weed Science,
44, 511-516.
·
Wiles, L.J., Wilkerson, G.G.,
Gold, H.J. y Coble, H.D. (1991). Weed
Science, 40, 546-553.
·
Zimdahl, R.L. (1980). International Plant Protection Center,
Oregon State University, 196 pp.